 m.quanpro.cn
m.quanpro.cn首页 > >
杂交骆驼知多少?
媒体:动物世界 作者:cuervo
专业号:钱佳弘
2024-01-22 09:13:09
众所周知,世界上有两种骆驼:单峰驼和双峰驼。肯定有不少人好奇,这两种骆驼之间能杂交吗?杂交后代长什么样?有几个驼峰?是否可育?今天我们就在下面这篇翻译的论文的汉语翻译版中一探究竟。
土耳其和哈萨克斯坦的双峰驼(Camelus dromedarius)和单峰驼(Camelus bactrianus)杂交畜牧实践:一次深度回顾
Dromedary (Camelus dromedarius) and Bactrian camel (Camelus bactrianus) crossbreeding husbandry practices in Turkey and Kazakhstan: An in-depth review
摘要
从古代起,在中东和中亚的许多地方,单峰驼和双峰驼之间的杂交已被进行。这一实践背后的主要推力是获得用于贸易车辆和军事行动的更好的驯服骆驼。今天,骆驼杂交只在两个地方被系统性进行:土耳其和哈萨克斯坦。在土耳其,骆驼杂交主要被进行用以获取一年一度的骆驼摔跤比赛的大型骆驼,而在哈萨克斯坦,骆驼杂交主要被进行用以获取更高产并更好抵抗中亚严酷气候的骆驼。
简介
单峰驼(Camelus bactrianus)和双峰驼(Camelus dromedarius)是通常发现于非洲、中东的两种不同和易于识别的家畜。两个物种占据完全不同的地理区域,其中单峰驼被发现于非洲北部、亚洲的中东和印度次大陆,而双峰驼仅被发现于亚洲内陆、中部和东部:中国、蒙古、哈萨克斯坦、吉尔吉斯斯坦、土库曼斯坦和阿富汗、伊朗北部、印度、巴基斯坦并直到土耳其东部(见图1)。
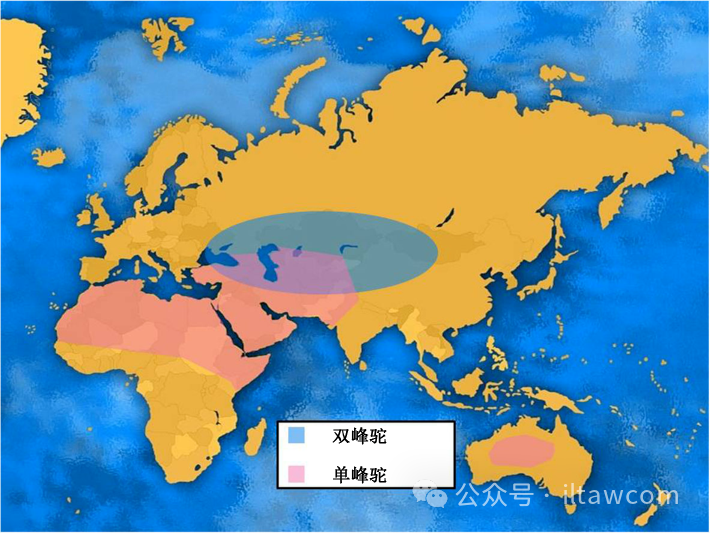
图1.单峰驼和双峰驼的分布范围以及重叠区域
当今的双峰驼分布范围远小于单峰驼,基本上局限在年均气温不超过21℃的地区(Mason 1979)。但事情并非一直如此。在历史上,双峰驼从公元前1000年到18世纪时的分布范围肯定更广,扩展到包括俄罗斯、中东大部分、土耳其、巴尔干和东欧的地区,由考古遗迹(Mukhareva 2007;Potts 2005;Peters和Driesch 1997, Pigière和Henrotay 2012)和诸如罗马硬币的普通物品上的双峰驼图像(见图2)所证实。

图2.来自公元98-117年罗马帝国阿拉伯行省,叙利亚-波斯特拉-德卡波利斯的罗马硬币
尽管双峰驼和在分类学上是不同物种,它们能够互相杂交。这产生出可育的F1(杂交第一代),多亏了杂交优势或来自亲本基因组间互动产生的优势,这些杂交后代有比它们的纯种亲本更好的生长率和生产性(Dzhumagulov 1980;Baimukanov 2002)。更重要的是,F1杂交后代能够更轻松地承受更冷、更潮湿的气候和崎岖地形。这些特性是对在两种骆驼间进行结构性的杂交畜牧实践的多个民族的发展的强有力激励。两种骆驼的杂交被假定起始于公元前1世纪,甚至更早时候,尤其是在两个物种重叠,或是在雌性单峰驼常见并且双峰驼能容易获取的地方:在中东、土耳其、从伊朗、伊拉克到阿拉伯南部,以及从土库曼斯坦到阿富汗北部(Bulliet 1975,2009;Potts 2004,2005;Uerpmannh 1999)。
单峰驼和双峰驼历史上的杂交畜牧实践
骆驼杂交的主要驱动力,是获取能够在有着巨大交易量和军事征战的地区承受崎岖地形和更冷、潮湿气候的强壮驯服骆驼的需要。这些地理区域包括现代的土耳其、巴尔干以及欧洲北部和东部。杂交骆驼出色的负载能力被反复记载(Bulliet 2009;Tapper 1985,2011),并且可能是杂交骆驼骨架残骸在多个拥有贸易中心和卫戍部队的考古地点被发现的原因。这些地点能在整个土耳其,以及欧洲东部和中部被发现(Pigière和Henrotay 2012;Vukovic和Bogdanovic 2013;Çakırlar和Berthon 2014;De Grossi Mazzorin 2006 2010;Bartosiewicz 1996;Bartosiewicz和Dirjec 2001;Galik等 2015)。奥斯曼军队因为杂交骆驼出色的承载力而广泛使用它们:400-500千克并能承受寒冷和多山的地形(Leese 1927;Burckhardt 1831;van Lennep 1870;Russell和Russell 1794)。在1529年奥斯曼大军发动的维也纳之围中,数千头骆驼被使用并在奥斯曼被击退后成为战利品告终。这些骆驼可能是16世纪由 Niklas Stӧn雕刻版画图像的基础,图上显示“土耳其人带着骆驼和单峰驼”,描绘一头有着两个靠近的非常小的驼峰的双峰驼,一定被描述为单峰驼和双峰驼间的杂交(见图3)。

图3.来自Niklas Stӧn在1530年创作的的“一个土耳其人带着一头骆驼和单峰驼”的陈旧版画,显示出单峰驼/双峰驼杂交种身上典型的单个锯齿状驼峰(承蒙德国 Landenmuseum-Gotha Herzogliches博物馆提供)。
根据17-20世纪间对在意大利San Rossore繁育的单峰驼成体和胎儿的描述(Lombardini 1879),即在主要驼峰前面的一个萎缩的驼峰,是骆驼杂交在欧洲相当普遍的进一步证明(Dioli 2014, 2018)。骆驼杂交通过雄性双峰驼和雌性单峰驼交配进行。这种操作已被考古发现证实,认定单峰驼是用于杂交育种的母本(Uerpmannh 1999)。这种杂交育种实践是几个作者报道最频繁的方式(Leonard 1894; Marsh 1856; Plassio 1912; Steel 1890),并被几个地区性旅行者提及(Burckhardt 1831;Olearius 1669)。这种杂交育种实践的确被使用的可视化证明在来自伊朗Haft Awrang的16世纪的手抄本装饰画中更加明显,其中一幅画显示一头雄性双峰驼和一头雌性单峰驼在一起,它们之间是杂交幼崽(Simpson 1997:92)。第一代杂交种F1骆驼的运载力和在多山地形与寒冷环境下的工作能力是常识。这在19世纪记录下美国进口单峰驼的报告中被提及(美国政府报告1857)。在那份报告中,对本次考察的负责的Lieutenant Porter甚至建议进口纯种雄性双峰驼,这样能和雌性单峰驼繁育F1杂交种(美国政府报告1857:123)。F1杂交种间进一步的自行杂交很少被进行,因为如此诞生的后代更小、更弱,并且“穷凶极恶”或无法应对恶劣天气(Burckhardt 1831; Fraser 1825; van Lennep 1870; Leonard 1894; Marsh 1856; Stewart 1881; Steel 1890)。历史上雄性双峰驼和雌性单峰驼间F1杂交种使用的另一个使用实在所谓的骆驼摔跤项目。伊斯兰艺术史学家创作的,描绘来自诸如阿富汗、印度西部和伊朗的广大地区的骆驼摔跤的多样中世纪微型绘画表明,这些项目相当普遍(Adamova和Rogers 2004)(见图4)。

图4.来自 Abd al Samad的绘画:两头打架的骆驼,大约在1590年。
在所有“骆驼摔跤”的历史绘画中,呈现的动物不是纯种的单峰驼或双峰驼,而是F1杂交种。单一的驼峰可以排除它们是双峰驼,而驼峰又过大过长,从而不可能是纯种单峰驼。此外,沿着脖子前侧、后背和头顶,以及前臂的浓密毛发排除这些动物是单峰驼,因为这些特征在纯种单峰驼中缺失,但在纯种双峰驼中一直提现。绘画中摔跤的骆驼显示口中有泡沫并且没有“吐出的红色肉球”或柔软上颚,这是在雄性单峰驼中存在但不存在于双峰驼和杂交种身上的器官。
现代单峰驼-双峰驼杂交生产
目前,骆驼杂交在土耳其东部被进行(Yilmaz和Ertugrul 2014),并且在前苏联共和国哈萨克斯坦以及邻国土库曼斯坦和乌兹别克斯坦以更结构化的方式被更广泛地进行(Faye和Konuspayeva 2012; Baimukanov等 2019; Dzhumagulov 1980;Ruchkina和Vakhitova 2008)。在土耳其,杂交生产的主要目的是获得为了每年冬季在该国西南部举行的“摔跤”项目的大型动物(Çakırlar和Berthon 2014; Yilmaz 2017; Yilmaz等 2015a, b, Yilmaz和Ertugrul 2014)(见图5)。在哈萨克斯坦和邻国,杂交的关注点是获得在产奶、驼绒和产肉方面更好,与此同时对中亚严酷气候更有适应性的骆驼(Imamura等 2017)。

图5.土耳其伊兹密尔2018年骆驼摔跤节上两头摔跤中的雄性F1杂交骆驼。注意嘴上的大量白沫并且边上的人准备在骆驼分出胜负时就将它们拉开。这可以阻止受伤。
来自土耳其的杂交生产方式
在土耳其,杂交生产旨在繁殖第一代杂交种F1,有时是第二代杂交种F2,更少见的情况是第三代杂交种F3。图6是目前在土耳其进行的单峰驼-双峰驼杂交实践的多种组合的示意图(Yilmaz和Ertuğrul 2015; Yilmaz等 2015a, b)。
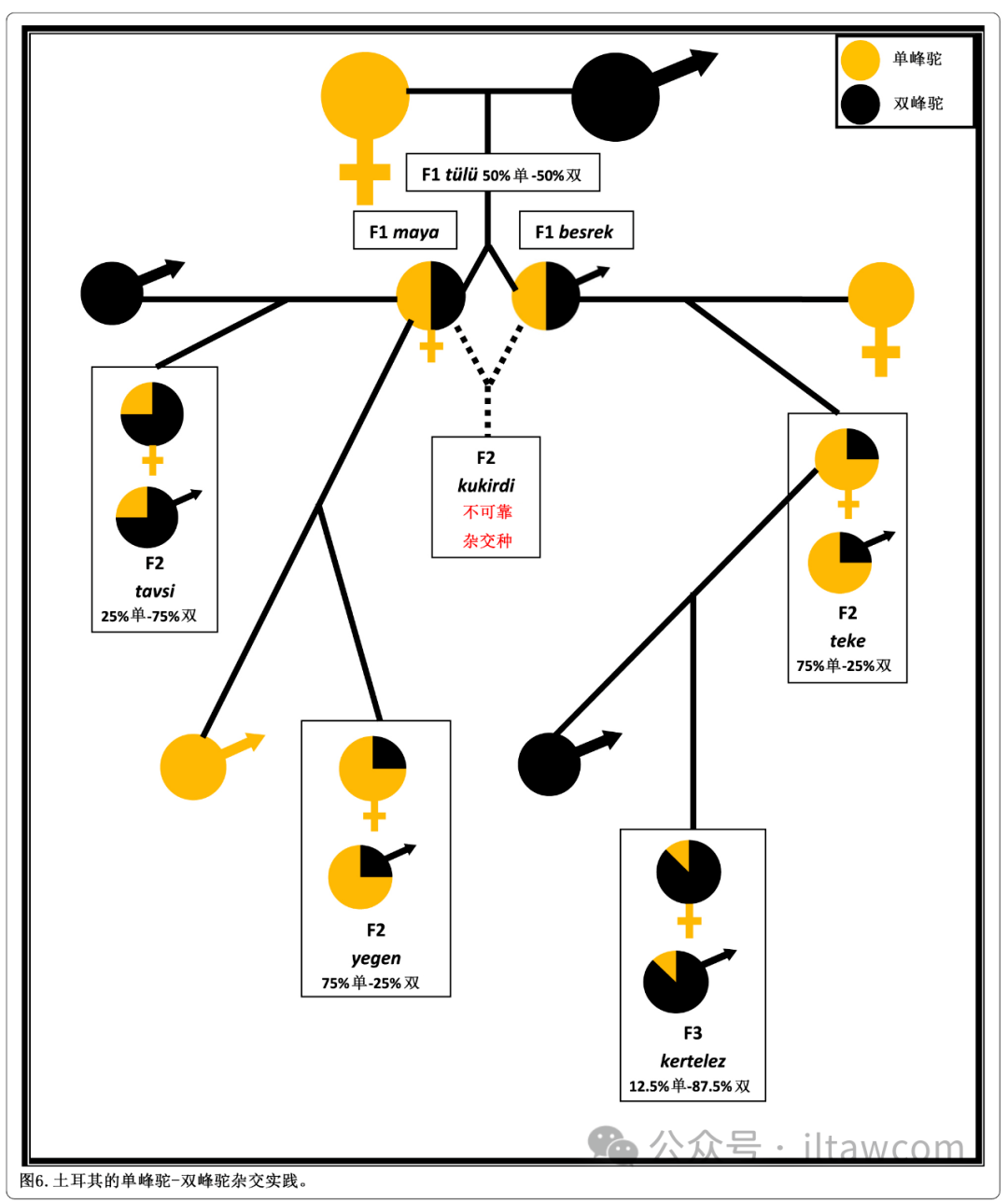
图6.土耳其的单峰驼-双峰驼杂交实践
土耳其杂交生产方式是基本的“双品种”杂交,由一头雄性双峰驼和一头雌性单峰驼交配组成(见图7)。在土耳其语中,这种F1的通用名叫tülü,更具体说雄性叫besrek而雌性叫maya(Yilmaz等 2015a, b)。这些杂交种外表上与单峰驼非常相似,尽管它们拥有一个从肩膀后就开始并延伸至超过腰椎的更大更长驼峰。这种特殊的形状被称为“杏仁峰”或者土耳其语的badem horguc(见图8)。

图7.一头雄性双峰驼和一头雌性单峰驼交配。注意雄性的前腿姿势舒适地跨在雌性身上并且如此一来给予雄性体位稳定性。

图8.一头土耳其F1杂交种tülü,显示出在土耳其语里叫做“杏仁峰”的典型拉长驼峰。
除了不寻常的加长外,F1杂交种的驼峰在前部有几厘米深的微小凹陷或缺口。这在营养好的短毛杂交骆驼身上清晰可见,在长毛杂交骆驼身上双手触诊时也可探测到。F1杂交种在脖子前侧、头顶和头后以及前臂上也有浓密毛发。F1间进一步杂交的F2没被进行,因为名为 kukirdi的杂交后代有着不合格的躯体发育(Dzhumagulov 1980),并且据说不良行为品质差到相当程度,以至于这些被描述为“对母亲和群体的威胁”(Çakırlar和Berthon 2014)。然而,进一步的杂交有时通过“回交”手段进行,第一代杂种的雄性和雌性与纯种单峰驼或双峰驼杂交以获取第二代杂交种(F2)。当雌性F1杂交种maya与雄性双峰驼杂交,后代被称为tavsi,而雄性F1杂交种tülü与雌性单峰驼杂交,后代被称为teke。它们不被用于摔跤,而是力畜,tavsi用于更冷的区域,而teke用于南部土耳其更热的区域。F1maya也可以和雄性单峰驼杂交产生名叫yegen的F2杂交骆驼;然而,这只是偶尔被进行,因其性能品质不被欣赏(Yilmaz等 2015a, b)。生产F3动物的进一步的杂交通过雌性teke与双峰驼回交进行;这种杂交种被称为kertelez,并且由于更高的双峰驼基因影响,它们适应土耳其更冷和多山的地区,例如托罗斯山脉。
来自哈萨克斯坦的杂交生产方式
中亚前苏联的最大国家哈萨克斯坦同时有单峰驼和双峰驼种群,拥有最复杂的骆驼杂交实践,以及俄罗斯和哈萨克斯坦科学家发展出的良好建立的杂交策略(Dzhumagulov 1980; Baimukanov 2002; Ruchkina和Vakhitova 2008)。存在许多地区性语言差异和方言;因此,鉴定杂交种的术语是复杂的(Bulliet 1975; Sala和Kartaeva 2017; Tapper 1985),并且在特定杂交种上容易混淆和不一致(Imamura等 2016)。本文遵从Baimukanov等(2019)采用的最新的杂交词源。两种主要的杂交手段被采用,都是基于双品种杂交开始,之后是多个回交步骤,并在图9中示意。
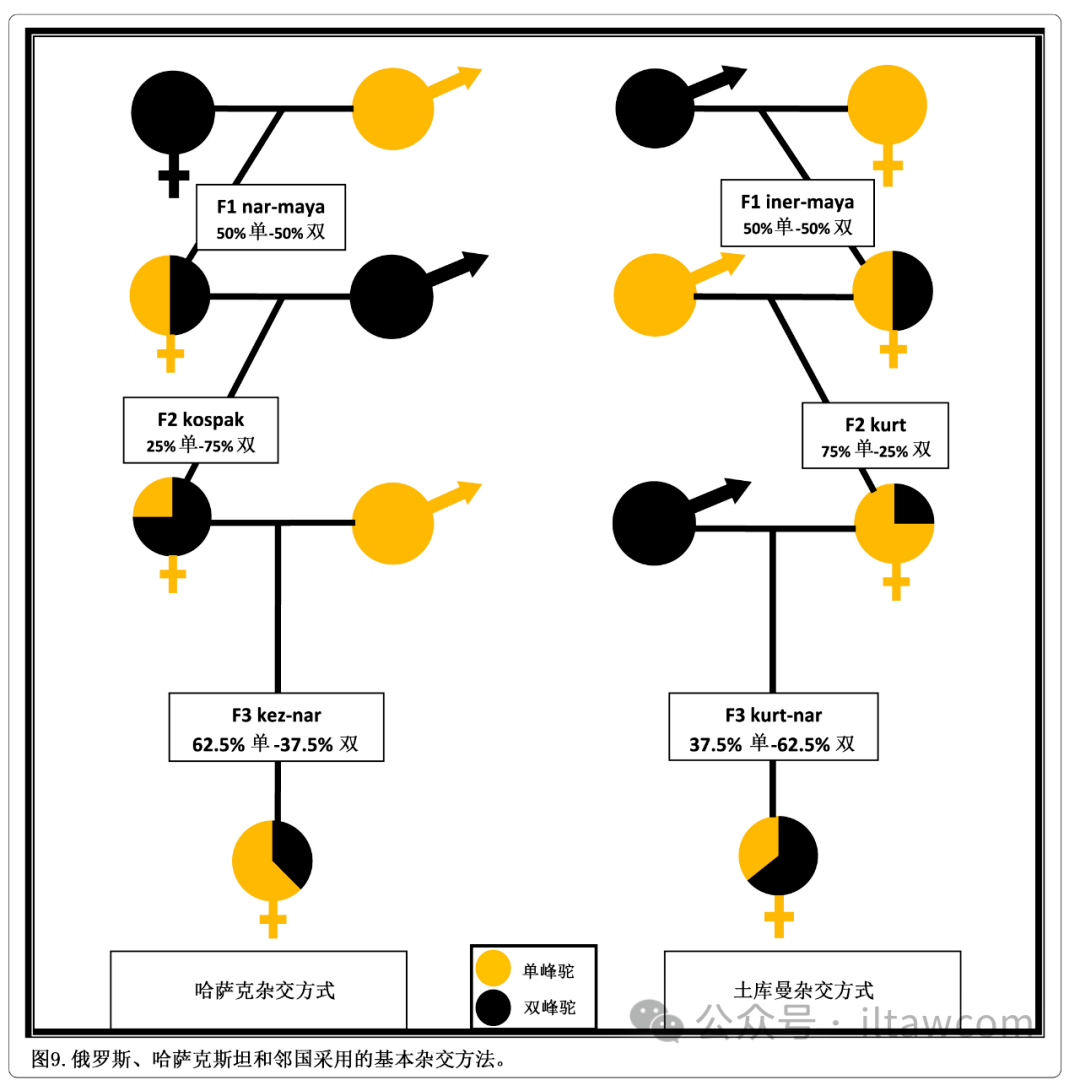
图9.俄罗斯、哈萨克斯坦和邻国采用的基本杂交方式。
所谓的“土库曼杂交方式”是指一头雄性双峰驼和一头雌性单峰驼杂交产生F1(与土耳其的方法一致),其雌性杂交后代叫iner-maya(等同于土耳其的maya)而雄性叫iner(等同于土耳其的besrek)。“哈萨克杂交方式”是指一头雄性单峰驼和一头雌性双峰驼杂交产生F1,其雌性杂交后代叫nar-maya而雄性叫nar(Baimukanov等 2018,2019)(见图10)。这种方式可能是因为哈萨克的单峰驼和土库曼的单峰驼或者arvana品种(见图11)拥有大而修长的身体结构,从而使得雄性克服在交配时雌性双峰驼后驼峰形成的阻碍。

图10.一头雄性单峰驼和一头雌性双峰驼交配。雌性的后峰和雄性的短体型迫使雄性表现出非常僵直的交配姿势。在这种情形下,交配的雄性单峰驼的前肢无法触地,而且雄性失去平衡并从雌性侧身滑下,导致无法完成交配。

图11.一头Arvana品种的雄性单峰驼种公驼。这种单峰驼品种体型大而修长,有一个拉长的驼峰和密集的棕色皮毛。该品种能忍受湿冷环境,并且有日均10-15升的良好产奶量。人高1.85米。
单峰驼和双峰驼都以蹲伏姿势交配;然而,当一头雌性双峰驼和一头雄性双峰驼交配时,其后驼峰迫使体型短的交配雄性单峰驼表现出一种非常僵直的不稳定姿势。在这种情形下,雄性经常从雌性侧面下滑并且无法完成交配。F1杂交种iner-maya和nar-maya,或者哈萨克斯坦克孜勒奥尔地区所说的bir-tygan,彼此间差异不大。两者都有单一的、从肩部延伸至腰部的极大驼峰,峰前有几厘米深的明显缺口。这些杂交种有比纯种单峰驼更多,也比纯种双峰驼相同身体部位处更多的毛发:颈部、头部、驼峰和前臂。在马的杂交案例中,母系的表型略微更明显(Lin等 2016)并且iner-maya杂交种倾向于更像单峰驼,而nar-maya继承更多的双峰驼特征,尤其是颈部和前臂的毛发(Ruchkina和Vakhitova 2008)。
雄性和雌性的F1杂交种之间不互相繁育,因为这样产生的后代会失去杂交优势(性状分离)。此外,它们的后代, 叫做jarbal或jarbay或dzharbai,拥有畸形胸部和关节的差的躯体发育,还有不可靠的脾气(Baimukanov 2002; Dzhumagulov 1980; Ruchkina和Vakhitova 2008)。F1杂交种所有的深层次杂交因此基于回交体系进行:雌性F1与其中一个原始亲本杂交,而雄性F1通常被去势(Ruchkina和Vakhitova 2008)。nar-maya(F1)雌性与雄性双峰驼的回交产生出名为kospak的F2杂交种,该杂交种的雌性后续与土库曼雄性单峰驼交配产生名为 kez-nar的F3杂交种群体。iner-maya(F1)雌性于雄性土库曼单峰驼回交产生名为kurt(等同于土耳其的yegen)的杂交种群体,该杂交种的雌性之后与雄性哈萨克双峰驼交配产生出叫做kurt-nar的F3杂交种群体(见图12和13)。

图12.杂交骆驼:a.kospak 3,b、c.kez-nar,和d.kurt

图13.杂交骆驼:a.iner-maya,b.nar-maya,c.kospak 1,和d.kospak 2
“土库曼杂交方式”和“哈萨克杂交方式”已知至少有数十年(Dzhumagulov 1980);然而,在哈萨克斯坦,对单峰驼和双峰驼取得更有利可图的利用的全新兴趣催生出更复杂的杂交方案的发展。
土库曼杂交方式的现代变化
图14总结出哈萨克科学家对土库曼杂交方式的多种改进。如在原版杂交方案中一样,雌性单峰驼与雄性双峰驼杂交后获得的iner-maya与雄性土库曼品种单峰驼回交,生出叫做kurt的F2杂交种。
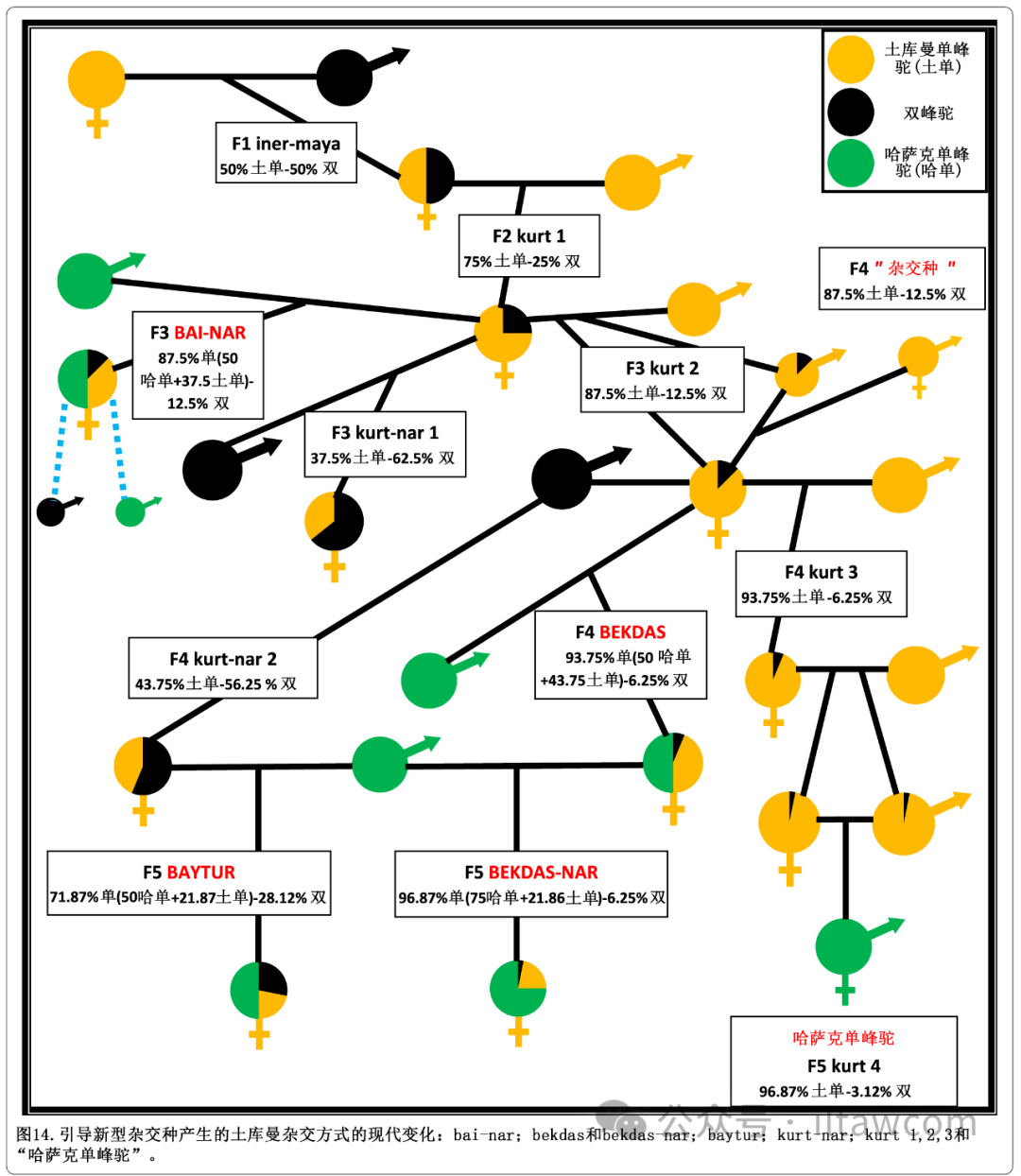
图14.引导新型杂交种产生的土库曼杂交方式的现代变化:bai-nar;bekdas和bekdas-nar;baytur;kurt-nar;kurt 1,2,3和“哈萨克单峰驼”。
在这之后,不同的杂交策略被实施。第一代的F2,kurt或kurt 1,再一次与土库曼品种单峰驼回交产出叫做kurt 2或 sapaldryks的F3杂交种,然后又和土库曼品种单峰驼回交产出叫做kurt 3的杂交种。该杂交种最终又回交生产出叫做kurt 4的F5杂交种。最后一代杂交种,有着96.8%的土库曼单峰驼基因和仅仅3.12%的双峰驼基因,之后通过群体内自行繁殖进一步发展,并创造出一个新品种,被称为哈萨克单峰驼(Baimukanov 2019; 哈萨克斯坦专利数据库 2003)。在表现型上,该品种能与别的单峰驼区分开,因为与土库曼单峰驼的三次连续回交已经稳定了它们单一驼峰的形状,并且移除了双峰驼在颈部、前臂和头部的典型体毛(见图15)。
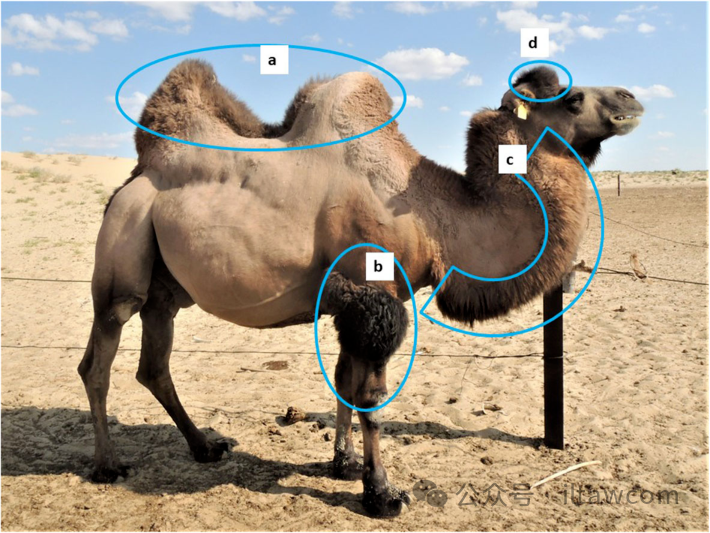
图15.双峰驼典型的表型特征:(a)两个驼峰的基部分开30-40厘米或是更多平坦区域;(b)浓密的前臂毛发;(c)局限在颈部前部边缘的浓密毛发,从喉部到脖颈基部;和(d)耳朵之间头顶部分的浓密毛发。这些区域的毛发生长在纯种单峰驼身上缺失,而在不同程度的杂交种身上总是有体现。
然而,哈萨克单峰驼的乳汁的脂肪含量比土库曼单峰驼的乳汁更高,哈萨克单峰驼的平均脂肪含量分别为4.5%(Baimukanov等 2019),而普通单峰驼的平均乳脂含量为3.52±0.74%(Zhao等 2015)。F3杂交种kurt 2也能种群间自行繁殖,产生被简单称作“杂交种”的商业性杂交后代。然而,雌性的kurt杂交种经常和纯种双峰驼回交。在这种情况下,F2杂交种kurt 1生下名为kurt-nar的F3杂交种,而F3杂交种产出名叫kurt-nar 2的F4杂交种(哈萨克斯坦专利数据库 2009a)。
一个三品种的杂交能通过F4杂交种kurt-nar 2和哈萨克单峰驼杂交产生出叫做baytur的新类型的F5杂交种(哈萨克斯坦专利数据库 2010c)。三品种杂交也可通过雌性F2杂交种kurt 1与雄性哈萨克单峰驼杂交以获得名为bai-nar的新类型F3杂交种(哈萨克斯坦专利数据库 2005)。bai-nar可以偶尔与新哈萨克单峰驼或纯种单峰驼杂交,产出被报道更早达到成熟期的杂交种。也能让一头F3杂交种kurt 2和哈萨克单峰驼再次杂交,获得名为bekdas的新型F4杂交种。最后,bekdas能与哈萨克单峰驼回交产生出名为bekdas-nar的新型F5杂交种(哈萨克斯坦专利数据库 2010d)。
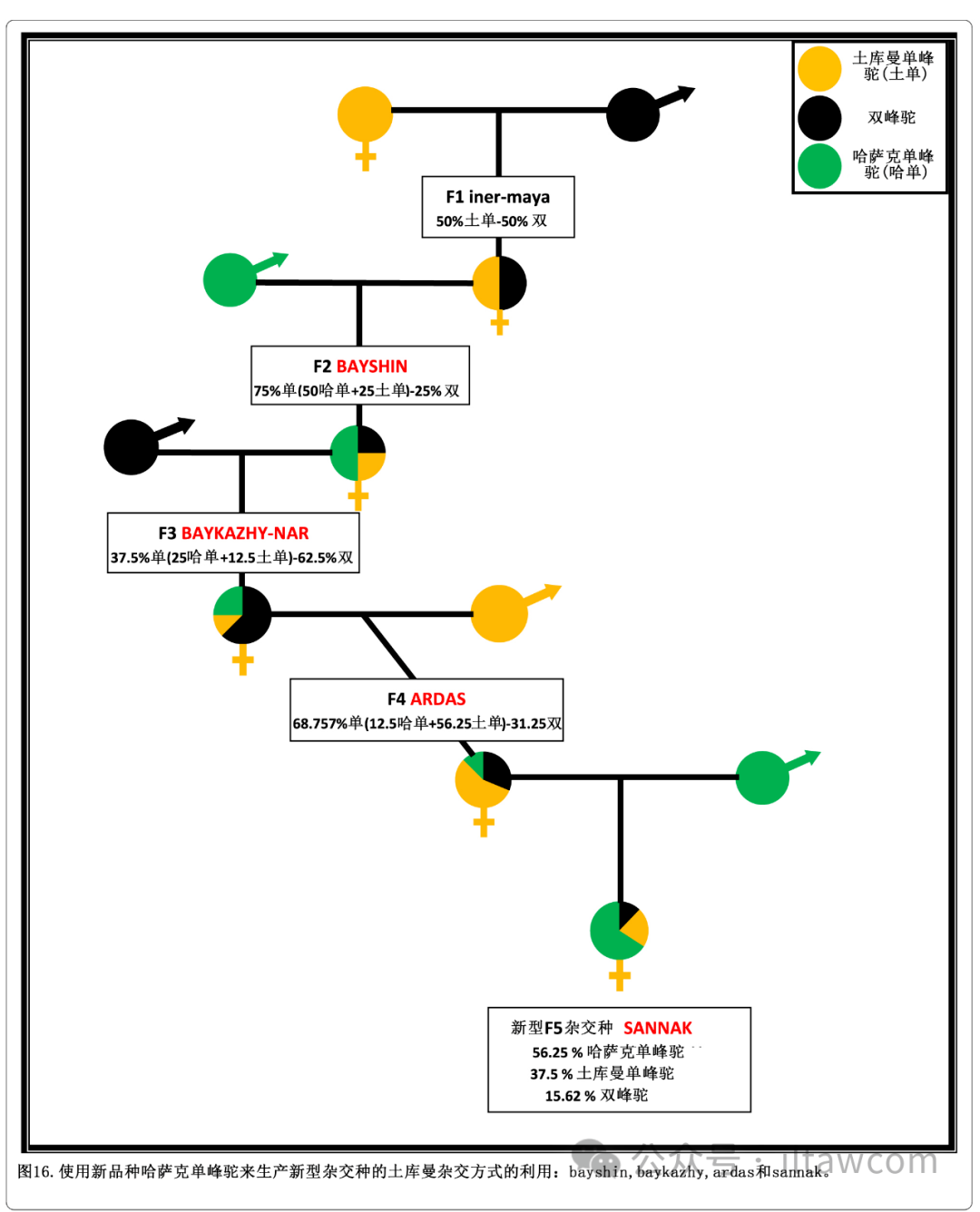
图16.使用新品种哈萨克单峰驼来生产新型杂交种的土库曼杂交方式的利用:bayshin,bakazhy,ardas和sannak。
在土库曼杂交方式中,使用到新品种哈萨克单峰驼的“三品种杂交”的进一步利用已经导致新杂交种的诞生。图16显示多种步骤。它以一头雄性新哈萨克单峰驼与一头雌性F1杂交种inter-maya杂交杂交产生新的名为bayshin的F2杂交种为开端。这在之后与纯种雄性双峰驼回交产生叫做baykazhy-nar的新型F3杂交种。该杂交种的雌性将进一步与雄性土库曼单峰驼回交产出名为ardas的新型F4杂交种,它之后又会和哈萨克单峰驼回交产出名为 sannak的新型F5杂交种(Baimukanov等 2016, 2017b, 2019)。
哈萨克杂交方式的现代变化
哈萨克杂交方式已经被哈萨克科学家深入修改。两个主要的繁育计划被制定,一个涉及到nar-maya和新品种哈萨克单峰驼杂交,另一个是使用土库曼单峰驼。图17示意表明了第一步。
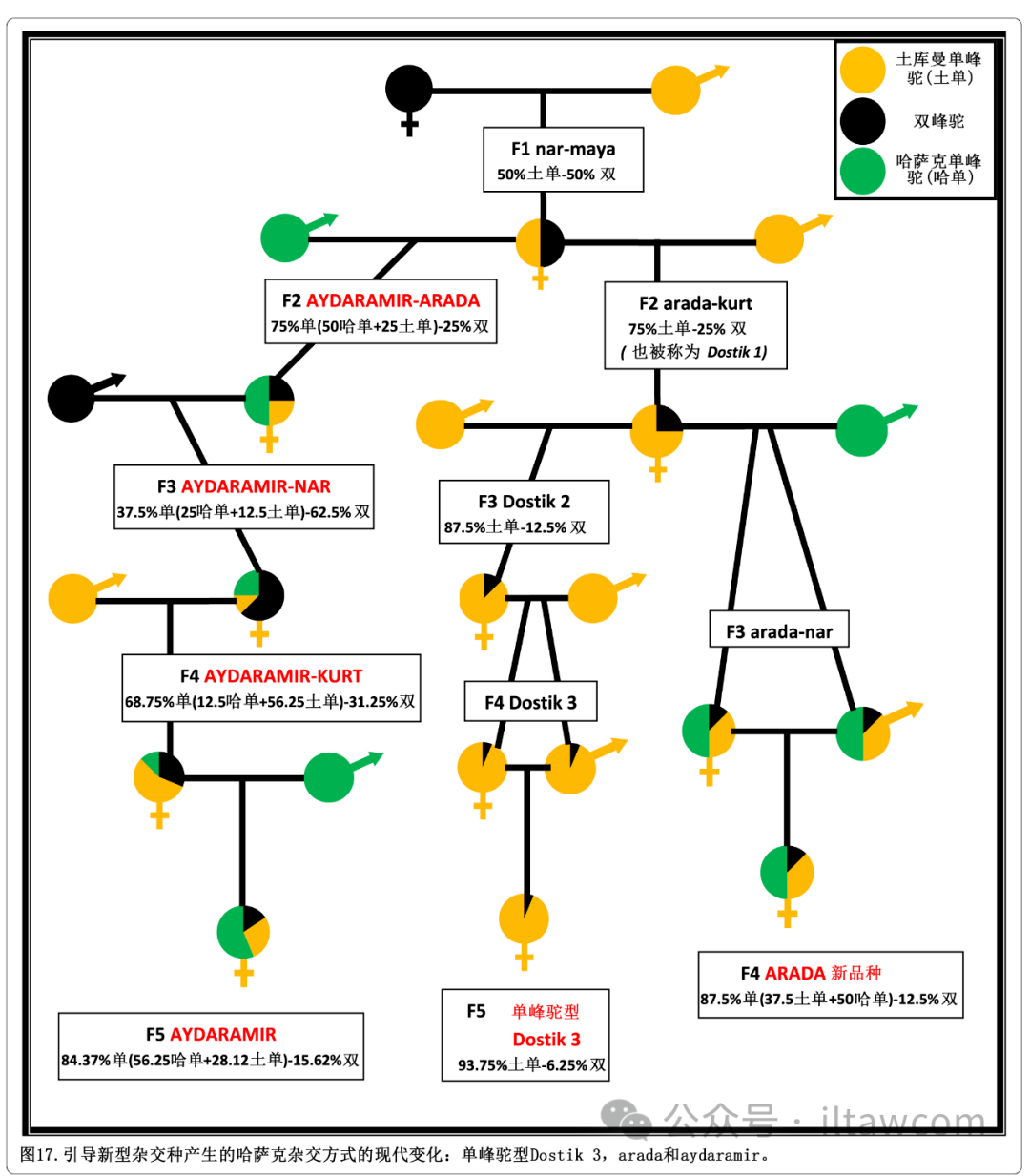
图17.引导新型杂交种产生的哈萨克杂交方式的现代变化:单峰驼型Dostik 3,arada和aydaramir。
这是一个使用新培育的哈萨克单峰驼的三种源杂交法。F1杂交种nar-maya和雄性哈萨克单峰驼回交生产出新的F2杂交种aydaramir-arada。该杂交种随后与双峰驼回交产生出叫做aydaramir-nar的F3杂交种,这种杂交后代再次与土库曼单峰驼回交产生出名为aydaramir-kurt的F4杂交种。这个最新的杂交种最终和新品种哈萨克单峰驼回交以获取叫做 aydaramir的新型F5杂交种(Baimukanov等 2019)。第二种杂交策略包括将F1杂交种nar-maya与雄性土库曼单峰驼杂交获取F2杂交种arada-kurt(也叫做Dostik 1)。该杂交种会和新品种哈萨克单峰驼杂交以获取名为arada-nar的F3杂交种。这些杂交种被进一步发展并允许自身内部繁殖,最终产生一种被认为全新品种的F4杂交种,名为arada。这个新品种有12.5%的单峰驼基因占比和87.5%的单峰驼基因遗传,其中土库曼单峰驼为37.5%,哈萨克单峰驼为50%(哈萨克斯坦专利数据库 2009b)。F2杂交种arada-kurt也可以通过与土库曼单峰驼持续回交来用于发展一系列叫做Dostik的杂交种。交配诞生出叫做Dostik 2的F3杂交种,随后是叫做Dostik 3的F4杂交种。这些最新的杂交种是相互之间繁育的,并且诞生被简单命名为Dostik 3类型的F5杂交种(Baimukanov等 2013; 哈萨克斯坦专利数据库 2014)。
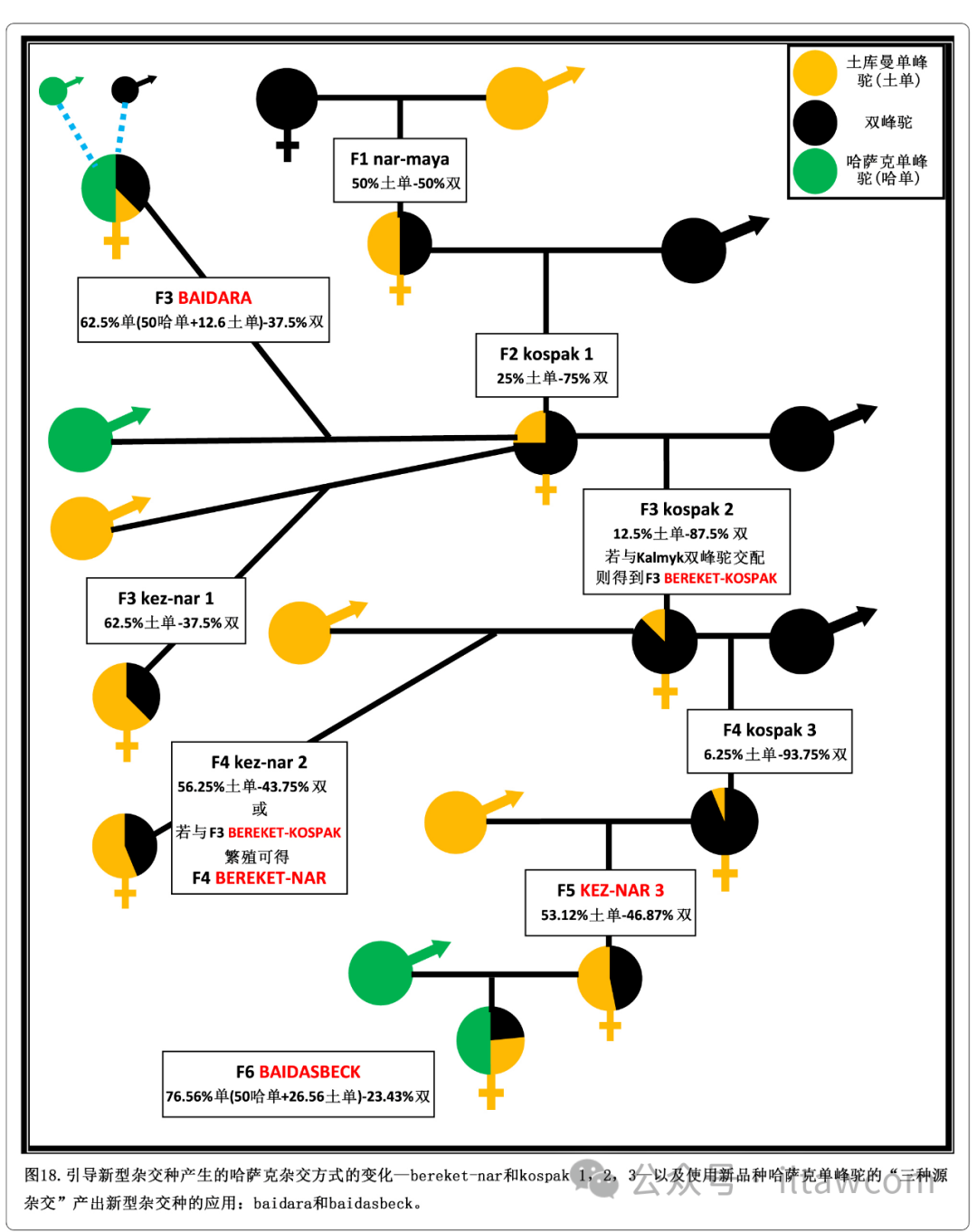
图18.引导新型杂交种产生的哈萨克杂交方式的变化—bereket-nar和kospak 1,2,3—以及使用新品种哈萨克单峰驼的“三种源杂交”产出新型杂交种的应用:baidara和baidasbeck。
哈萨克斯坦杂交方式的第二处变化在图18中展示(Baimukanov等 2019; 哈萨克斯坦专利数据库 2008a,2009c,2010a, b)。有了这个繁殖方法,F1杂交种nar-maya与雄性双峰驼回交产出叫做kospak 1或bal-kospak的F2杂交种。该杂交种之后与雄性双峰驼回交产出叫做kospak 2或myrza kospak的F3杂交种,又再一次获得叫做kospak 3或nar-kospak的F4杂交种(哈萨克斯坦专利数据库 2008b)。所有的kospak杂交种能与土库曼单峰驼回交产出名为kez-nar的多种类型杂交种。F2杂交种kospak 1产出F3杂交种kez-nar 1,F3杂交种kospak 2产出F4杂交种kez-nar 2,F4杂交种kospak 3产出F5杂交种kez-nar 3(哈萨克斯坦专利数据库 2008a)。
一个对此种杂交方案的重要补充包括引入“三品种杂交策略”,让F2杂交种kospak和哈萨克单峰驼杂交并以此产出全新的名为baidara的F3杂交种(哈萨克斯坦专利数据库 2009c)。baidara有时可以与哈萨克单峰驼或纯种双峰驼杂交,因为这似乎增加了早期的生殖成熟度。哈萨克单峰驼也被利用,通过与雌性F5杂交种kez-nar 3杂交产出名为baidasbeck的新的F6杂交种(哈萨克斯坦专利数据库 2010b)。最近,该方案的一个变动已经被添加,是通过使用大体型的双峰驼:Kalmyk品种(见图19)。

图19.Kalmyk品种双峰驼种公驼。这种双峰驼是巨大的,活体重量可以超过1200千克。该品种能生产6-9千克的好驼绒并且日均产奶量为2.5-3.5升。人高1.85米。
当雌性F3杂交种kospak与Kalmyk品种的雄性双峰驼杂交时,得到一个名叫bereket-kospak的珍贵杂交后代,该杂交种与雄性土库曼单峰驼杂交时产生名为bereket-nar的新型F4杂交种(哈萨克斯坦专利数据库 2010a)。
主要的已确立的杂交优势的概述
哈萨克和俄罗斯科学家发展的单峰驼和双峰驼杂交繁育实践的主要目的,是培育出拥有比单峰驼跟高的产奶能力,以及能够在基本放养的情况下在中亚严酷气候下生存的骆驼品种。这些地方的气候条件以炎热的夏天和大雪封山的季度零下的冬天为特征。大量的拥有这些品质的杂交种被培育。尤其有趣的是bainar, baykazhy, aydaramir-arada, aydaramir-nar,
baidasbeck和baidar杂交种,它们有着比土库曼单峰驼更高的产奶量。而F2杂交种:bayshin和aidaramyr-nar、所有的F4杂交种(除去kurt 3)和F5杂交种sannak和aidaramyr有着比单峰驼和双峰驼更高的活体重量。俄罗斯和哈萨克研究者的工作(Baimukanov等 2016, 2017a, b, c, 2018, 2019; Dzhumagulov 1980)已经建立起如下关于骆驼杂交实践的通常规则:
· 雄性杂交种不被推荐(除了F5 kurt 4, F4 Dostik 3, F3 arada-nar)。
· F1杂交种的体型比单峰驼和双峰驼任一纯种亲本大5-12%,并有更好的工作能力和更高忍耐力。用哈萨克杂交方式获得的F1杂交种比土库曼杂交方式获得的更大。杂交优势的积极影响从1岁起更大。
· 所有的杂交种有着比单峰驼和双峰驼更长的生产寿命,其中F1杂交种nar-maya和F3杂交种kez-nar在生命周期中分别能泌乳7.3次和6.8次。
· 当F1杂交种内部自己繁殖时,杂交优势的积极效果完全不见,并且产生的名为jarbay的F2杂交种是一种不合格的动物。
· 杂交驼崽有着比单峰驼幼崽(平均出生体重34.5±0.09千克;Nagy和Juhász 2019)和双峰驼幼崽(平均出生体重34.55±7.17千克;Zhao等 2000)更高的出生体重(F1平均出生体重 45.4±0.842.1千克)。它们的生长率也更高。
· 杂交种身上单峰驼基因比例越高,就增加总体产奶量,这是由于:更长和更充足的泌乳期;增大的乳房尺寸和结构,包括更好的四部分对称性和发育;增加的乳头间隔、尺寸和向下的方向。然而,这也减少奶中的脂肪比例,削弱驼绒产量、早期性成熟以及对更冷气候的适应性。
· 杂交种身上双峰驼基因比例越高,就增加驼绒产量、活重和奶中的脂肪比例,但是削减泌乳期时长和绝对产奶量。孕期也延长。
· 所有的杂交种的妊娠期都比双峰驼短(平均442.5±5.1天),但比土库曼单峰驼(平均425±3.9天)和哈萨克单峰驼(平均416.2±3.1天)长。单峰驼基因缩短了孕期。F4杂交种拥有最接近土库曼和哈萨克单峰驼的妊娠期。
· 所有杂交种的驼绒产量都比双峰驼低。
· 所有杂交种的乳脂比例低于双峰驼(平均5.34±0.60),但高于纯种单峰驼(平均3.52±0.74)(Zhao等 2015)。
总结和建议
对于目前只饲养双峰驼的中亚地区,骆驼杂交可能在提高杂骆驼生产力方面起到重要作用,因为单峰驼无法在中亚气候下生存。的确,利用杂交种来提高驼奶产量的倾向在哈萨克斯坦的一些地方早已是可观测的(Imamura等 2017).在哈萨克斯坦很多地方,单峰驼、双峰驼和它们的杂交种是自由放养并自然交配的(见图20)。

图20.a,b.未知代数的杂交骆驼,可能是F4-F5。c.杂交骆驼:从左到右为bereket、kospak和iner-maya。d.杂交骆驼:从左到右为kez-nar、kospak和kez-nar 3。
在这种情况下,无人管理的交配可能发生并且未被记录,产生出带有未知单峰驼/双峰驼基因组成的动物。所有这样的杂交种,尽管形态上与纯种单峰驼一致,仍然携带可能影响杂交种生产力的双峰驼基因。基于表型特征的杂交骆驼鉴定是不可靠的,尤其是对诸如F4-F6的后阶段杂交种(表1和图15)。

表1.提及单峰驼-双峰驼杂交骆驼的表型双峰驼特征的列表。
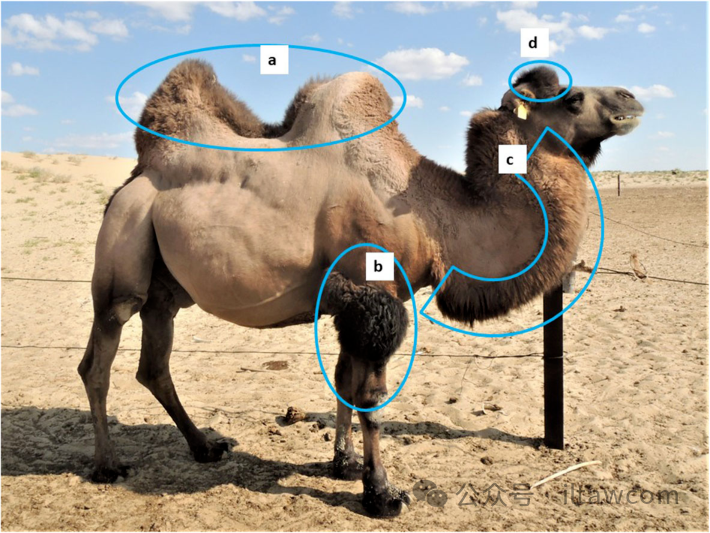
图15
因此,严格的谱系记录,结合上永久的骆驼鉴定,是清晰鉴定杂交个体,以及单峰驼与双峰驼在其基因组成中确切基因比例的更好工具。先进基因技术和亲子鉴定增加的可利用性将允许更好地理解单峰驼、双峰驼和它们的杂交种,可能允许检测那些与增加生产力和适应性相关的宝贵基因特征(Burger等 2019)。
补充材料



同为咸海一带的骆驼群。图中可以清晰地看到表型为双峰驼、单峰驼的个体,以及有两个驼峰但分界不明显的杂交骆驼。
值得一提的是,新疆过去也曾有过单峰驼饲养,但是由于数量稀少,加上当时缺乏保育观念,导致中国境内的单峰驼淹没在双峰驼的洪流之中。以下仅由黑点组成的和田单峰驼的照片是国内单峰驼唯一留存下来的痕迹。


图中的骆驼可以看出有一个驼峰,但脖颈和头顶毛发浓密,有可能也是杂交骆驼。
阅读
1487
| 版权声明: 1.依据《服务条款》,本网页发布的原创作品,版权归发布者(即注册用户)所有;本网页发布的转载作品,由发布者按照互联网精神进行分享,遵守相关法律法规,无商业获利行为,无版权纠纷。 2.本网页是第三方信息存储空间,阿酷公司是网络服务提供者,服务对象为注册用户。该项服务免费,阿酷公司不向注册用户收取任何费用。 名称:阿酷(北京)科技发展有限公司 联系人:李女士,QQ468780427 网络地址:www.arkoo.com 3.本网页参与各方的所有行为,完全遵守《信息网络传播权保护条例》。如有侵权行为,请权利人通知阿酷公司,阿酷公司将根据本条例第二十二条规定删除侵权作品。 |